Apró cseppek indíthatták útjára a földi élet fejlődését
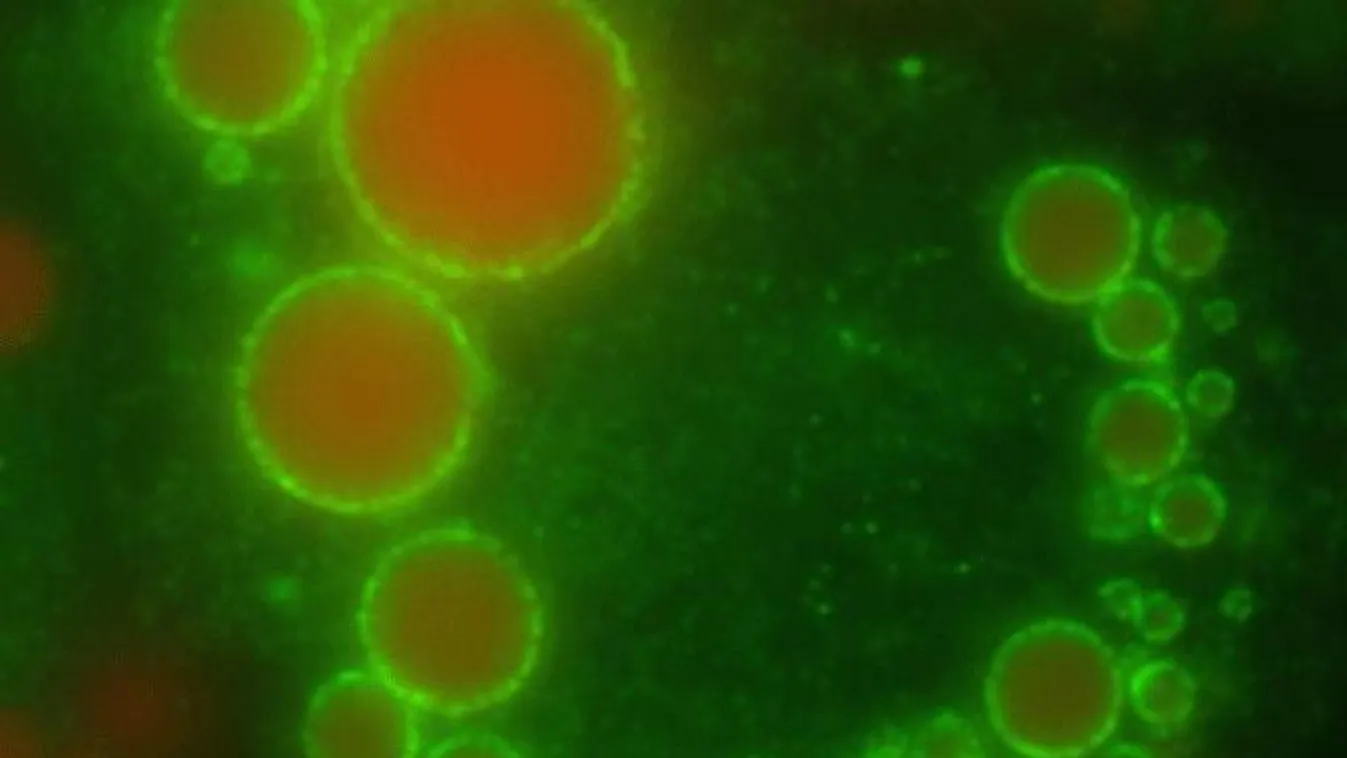
Lipidcseppek mikroszkóp alatt
Nézze meg a sporthíreket is
Nézze meg a sporthíreket is
Az első élőlényekről csak annyit tudunk, hogy kémiai rendszerek lehettek, melyek képesek voltak valahogy létrehozni saját maguk másolatát, miközben felhasználták a közelben elérhető egyszerű molekulákat. Ilyen, önmaguk sokszorosítására képes kémiai rendszerek ismertek, azonban a világukban farkastörvények uralkodnak: amelyik gyorsabban másolja magát, az kerül többségbe.
A félelmetesen hatékony molekuláris másológépek vagy más néven replikátorok világa lehetett ez – nehéz elképzelni, hogy milyen út vezethetett innen az élet ma ismert bámulatos komplexitásához. A másoláson túlmutató funkciók valószínűleg kikoptak, hiszen a többletfunkció egyúttal hosszabb molekuláris kódot jelentett, a hosszabb kód másolása pedig hosszabb időt vett igénybe, így a funkciót nem hordozó másológépek egyszerűen túlszaporodták őket.

Lehettek persze olyan meglepő módokon változó környezeti tényezők, amelyek egyre bonyolultabb molekulaszerkezetek kialakulását követelték meg, azonban van egy ennél egyszerűbb, a természetben sokszor felbukkanó lehetőség a komplexitás növelésére:
a kompartmentizáció,
melynek során az élőlények valamilyen okból környezetüktől elkülönülő csoportba tömörülnek, és a szelekció erre a csoportra hat.
Ha ilyenkor az egyes résztvevők tudnak olyasvalamit tenni a csoportért, ami javítja a túlélési esélyeit, ez a tulajdonság megmaradhat. Ekkor már nem a brutálisan hatékony, de a közösség számára értéktelen másológépek állnak nyerésre, hiszen amelyik közösségben felszaporodnak, az egy idő után hátrányba kerül, a többi, többségében „becsületes" tagokból álló közösséggel szemben.
Ha az iskolai osztályban gesztenyét gyűjtenek, és mindenkit megkérnek, hogy csak borítsa be, amit hozott a terem végében egy nagy ládába, előnyben lesznek a bliccelők – de a helyzet nyomban egész más megvilágításba kerül, ha csak azok az osztályok mehetnek el az év végi hatalmas buliba, ahol elég gesztenye gyűlt össze.
Az evolúcióbiológia ismeri ezt az állapotot – az ilyen kompartmentekbe tömörült replikátorok lennének sejtjeink korai elődei, a protosejtek. Van azonban velük egy kis gond. Hogyan jön létre maga a kompartment? A csoportot körülölelő burok felépítésének receptjét magukban kell hordozniuk a benne működő replikátoroknak. Ez a recept pedig, akármi legyen is, információ, vagyis másolása többletidőt vesz igénybe.
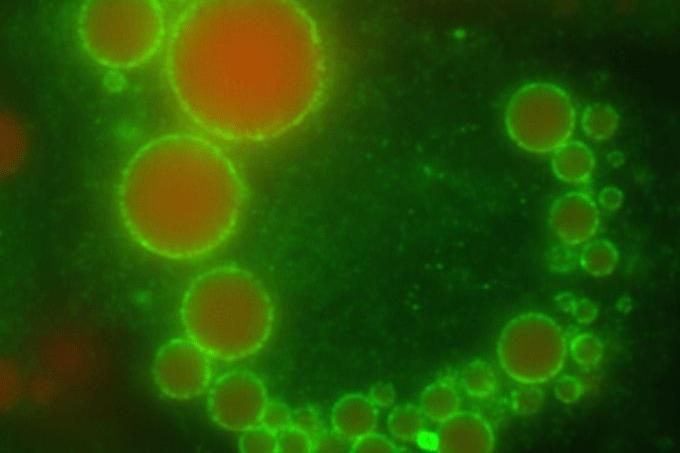
Ismét visszajutottunk a kiindulóponthoz: az önmaguk másolására optimalizált molekuláris gépek nem tűrnek meg felesleges információt – hogy jutunk hát el egy teljes kompartmentreceptig?
Szathmáry Eörs és kutatótársai a kérdést megkerülve, más oldalról közelítették meg a problémát: ha a replikátorok maguk nem képesek előállítani a kompartmenteket, kialakulhatnak-e esetleg más úton? Eredményeikről a Science magazinban tették közzé.
Nos, nem kell túl messzire mennünk, hogy lássuk: igen, kialakulhatnak! Elég csak kisétálnunk a konyhába, és már a mosogatóban megtalálhatjuk a megoldást.
A serpenyő öblítővizében szétváló olajcseppek vagy a csöpögő csap által felvert vízcseppek éppen ilyen apró kompartmenteket adnak,
melyek, ha ideiglenesen is, de elválasztják környezetüktől a belsejükben található anyagokat.
Ilyen és ehhez hasonló átmeneti kompartmentképző folyamatokkal minden bizonnyal a négymilliárd évvel ezelőtti Földön is találkozhattunk volna. Most már csak az a kérdés, hogy ezek segítenek-e a replikátorok problémájának megoldásában. A kutatók nekigyűrkőztek, és megszületett a kísérleti rendszer.
A természetben ma működő legegyszerűbb replikátorok a vírusok, melyek ugyan nem képesek teljesen önállóan másolni magukat, azonban fondorlatos módokon rá tudják venni a sejteket arra, hogy dolgozzanak helyettük. Működésük viszont a sejtek rendkívül összetett rendszerében zajlik, így eredeti formájukban nem alkalmasak az ősi replikátorok modellezésére.

A kutatók ezért addig csupaszították egy RNS-vírus másolási mechanizmusát, amíg tényleg csak a legszükségesebb elemek maradtak meg belőle: az örökítőanyag és egy replikáz, vagyis másolóenzim. Ha ezeket – no meg az RNS építőköveit, a nukleotid-trifoszfátokat – összekeverik egy oldatban, a replikáz példányai dolgozni kezdenek, és addig másolgatják az oldatban jelen levő RNS-láncokat, amíg az építőkövek el nem fogynak.
A kiválasztott vírus replikáza azonban nem tud bármit másolni – csak olyan RNS-szálakkal foglalkozik, amelyek két vége összetapadva kiad egy meghatározott kulcsot. Ha ez a kulcs jelen van, bármilyen RNS-szakasz behelyezhető a kulcs két fele közé, és a replikáz lemásolja. De mi kerüljön a kulcs két része közé?
Nos, a kutatók nem véletlenül választottak RNS örökítőanyagot, hiszen a ribonukleinsavak némelyike képes a jól ismert fehérjeenzimekhez hasonlóan kémiai reakciókat katalizálni – ezeket hívjuk ribozimeknek. A kulcs két részlete közé egy ilyen ribozimet illesztettek, mely lényegében molekuláris ollóként viselkedik, és meghatározott ponton képes elvágni egy RNS-molekulát, amely a közelébe kerül. E molekuláris olló jól példázza azt a másoláshoz nem kapcsolódó funkciót, mely növelhetné kis replikátorunk komplexitását.
Könnyen látható, hogy a
míg az olló nem járul hozzá valahogy a replikátor sikeréhez, hamar kikopik az utódok kódjából,
hiszen azok a másolási hibák nyomán keletkező mutánsok, melyek kódja rövidebb, gyorsabban másolódnak, így hamar többségbe kerülnek. Iskolai példánkkal: ha senki nem nézi, ki mennyi gesztenyét önt a ládába, és úgyis mindenki elmehet a buliba, előbb-utóbb drasztikusan megnő a bliccelők száma.
Szathmáry Eörsék kísérletükben a molekuláris ollónak vágnivalót is adtak: egy különleges molekulát ragasztottak egy rövid RNS-szakasz végére, éppen úgy, hogy az olló képes legyen levágni. A ráragasztott molekula különlegessége, hogy ebben a levágott, önálló formájában narancsszínben fluoreszkál. Ezzel a módszerrel gyorsan és könnyedén megmérhető, mennyi olyan replikátor van az oldatban, amelyben működik a molekuláris olló.
A kutatók ezután kijelölték a példabeli iskolai osztályokat, és meghirdették az év végi bulit. Készítettek egy olyan gépet, mely apró cseppekre bontotta a kiindulási oldatot, majd megvárta, hogy a cseppekben kedvükre másolgassanak a vírusreplikázok, végül aszerint tartotta meg vagy dobta ki a cseppeket, hogy elegendő számú molekuláris olló működött-e bennük, vagyis kellően erős narancs fluoreszcenciát mutattak-e. A megtartott cseppeket összeöntötték, és az ebben az oldatban található replikátorokkal újrakezdték a kört.
A másolás természetesen hibákkal járt, és megjelentek olyan mutánsok, amelyeknél nem működött az olló, viszont gyorsabban másolták őket a vírusreplikázok. Ezeket a mutánsokat most már parazitáknak tekinthetjük, hiszen az ő működésük nem tesz hozzá semmit az őket tartalmazó kompartment sikeréhez. Amikor a kísérletet a narancs fluoreszcenciára nem szelektálva végezték, az eredmény a vártaknak megfelelően alakult: az ollót tartalmazó RNS-szakaszok és velük a narancs fluoreszcencia a 7. kör után eltűnt.
Nem mellesleg érdemes megjegyeznünk, hogy a „felesleges" kód eltűnése itt lassabb volt, mint ha egyszerűen összeöntötték a rendszert egy közös oldatba. Vagyis az ideiglenes kompartmentek puszta létükkel, szelekciós hatás nélkül is adnak némi esélyt a „kis hibás" replikátorok túlélésének.
Amikor azonban bedobták az év végi bulit, vagyis csak a narancs fluoreszcenciát mutató cseppeket engedték át a következő körbe, még a 9. körben is jelentős mennyiségben voltak a működő ollót tartalmazó, replikálódó RNS-szakaszok. A kísérleteket egy matematikai modellel jóval több körre kiterjesztették, és különféle másolási sebességű parazitákra vizsgálták. A számítások szerint az efféle rendszerekben végül beáll az egyensúly, és nem tűnnek el az ollóval rendelkező replikátorok.
Ezzel bebizonyították, hogy ha létezik olyan természetes mechanizmus, amely rendszeresen ideiglenes kompartmenteket hoz létre, és egy másik körülmény ezeket a kompartmenteket valamilyen tulajdonságuk alapján szelektálja, akkor az segíthet abban, hogy
a replikátorok a brutálisan hatékony másológép mellett valamilyen egyéb, a másoláshoz közvetlenül nem kapcsolódó funkció kódját is megtartsák.
A kísérletnek azonban volt egy másik, hasonlóan izgalmas eredménye. A rengeteg mutáns között mindig vannak nagyon gyorsan másolódó „kemény" paraziták és kevésbé gyorsan (de az ollót tartalmazó példányoknál azért gyorsabban) másolódó „lágy" paraziták. A modellszámítások azt mutatták, hogy ennek a rendszernek utóbbiak a nagy nyertesei. A szelekciós folyamat mindaddig, míg a funkció (az olló) elegendő mértékben érvényesül, megtűri a lágy paraziták jelenlétét, ami nagyobb információs változatosságot enged az ideiglenes kompartmentek replikátorainak. A nagyobb változatosság pedig egyúttal jobb alkalmazkodóképességet is jelent – egyfajta „információs bank" jön létre, melyből kiindulva sokféle fejlődési út lehetséges.
Gondoljuk csak meg: ha létezik egy hatékonyabb molekuláris olló, amely sok apró mutációs lépésben érhető el az eredetiből, akkor a rendszerben meglevő kis „lazaság", ami megengedi a hibás példányok fennmaradását, segíthet végighaladni ezen a lépéssorozaton. Nem is beszélve arról, hogy a „hibás" példányok között megjelenhet egy teljesen új funkció is, ami máshogy járul hozzá a csoport sikeréhez.
A cikk kicsit bővebb változata az MTA oldalán olvasható.












